



























既然碘在自然界中并不富裕,那么为什么甲状腺激素一定要用碘来合成?
你是《自然选择》项目组中的一员。 你接到上级指派的新需求:给脊椎动物的屎山代码里加个新系统,要能够控制机体消耗能量、制造蛋白的速度、还要能调节现有的各个系统对各种激素的敏感性,对身体、智力的发育,代谢都要有重要影响。 最高级别响应!! 你看着这闹着玩似的需求陷入沉默,这牵一发动全身的狗屎代码微调都常常跑不动,现在竟要加个 overrule 的管理员级别应用??程序员的命也是命啊。但牛马精神还是驱使你点开了素材库,开始思索怎么用相对简单的方式把以上所有功能加于一个模块上。 既然这个模块的权限那么高,就先把它设定为最大的内分泌腺吧,你思索到。 大致方向有了,那具体的信号传递方式呢? 细胞上的受体容易加,但传递信号的分子最好单一。 因为假如分子多样,那受体也要跟着多样,产生信号分子的器官的复杂性就会提升,信号传递过程中的鲁棒性降低,容易受到屎山系统的干扰而故障。 另外,代谢这个信号分子的通路也最好单独设计,不能是体内广泛存在的酶,不然出 bug 是迟早的事。 但是常规的由 C,H,O,N 组成的分子也容易出 bug。 项目组曾经的战绩是用简单的链烯烃通过一步串联反应产生四五根 C-C 键,构建出了高度刚性、立体专一的甾体和三萜化合物。这神来一笔瞬间提升了分子的复杂度和稳定性,和其他稀稀拉拉、扭来扭去的分子拉开身位。此类骨架用作信号分子(激素),后期只需做些许微调(衍生化),就能满足差异化和不同需求。如此一来,对应的蛋白受体设计起来也省心不少,不需要大幅修改。 尽管如此,甾体前期的物料准备和后期的衍生化还需要十几二十多步的转化,这还没算上代谢清除过程,简直费时费力。对于调控代谢水平这种即时性很强的需求不能充分的满足。 更要命的是,假如为新模块设计的这个信号分子难以被代谢清除而积蓄,或者受体和其他长得差不多的分子结合,那用户端代谢水平就会不断增加,长此以往早衰 go die。 反之,如果这个由 C,H,O,N 组成的信号分子被广泛存在的氧化还原酶 / 水解酶等等玩意儿截胡,被迫参与了一些针对 O,N 基团的反应,那可能还没来得及传递信号就丧失了功能,用户端代谢水平于是下降,免疫调动不起来,长此以往也 go die。 因此,你开始考虑添加一套完全不同于现有生化反应的新的、能快速反馈和调节的化学反应,正交(互不干扰)的化学不容易出 bug,所以最好加个新的元素,围绕这个元素产生的反应必然和现有体系正交,配得上这个模块管理员级别的地位。 于是在素材库里,你企图为 C,H,O,N 做骨架的小分子增添一些新的化学元素。 元素周期表 你瞅了眼碱金属(Li, Na, K...)和碱土金属(Ca, Mg..),之前已经被设计组用作离子通道了,pass。 过渡金属(Fe, Zn, Mo, Cu..)多被设计在蛋白里,做信号小分子很难,还有代谢和积蓄的问题,pass。 类金属(B, Si, Ge, As...)容易被氧化还原,而且碳与类金属的化学键很难在复杂的生理环境中稳定。 稀有气体(He, Ne, Ar... )化学惰性,第六七周期的元素基本都是金属,难获取还有放射性更不用考虑。 所以你将目光聚集在非金属上,除了 C,H,O,N 外,还剩 F,S,P,Cl,Se,Br 和 I,碳基生物能和这些元素很好的兼容,正因为能形成稳定的碳 - 杂原子键。 C-F 键能强,难代谢,更难在体内合成,游离的氟源(F2)在自然界中不存在,即便摄入氟离子,也优先和体内的钙离子结合沉积,pass。 S 已经被开发在启动子甲硫氨酸和双硫键里,也就是可以形成蛋白高级构象的半胱氨酸里,还有高能的硫酯键,用在了诸如补体系统的 C3b 里。 如果代谢这个信号分子涉及改变硫元素的氧化态,势必需要氧化还原酶,而这个酶未必不能作用在半胱氨酸上。此外,生理条件下还原型谷胱甘肽(GSH)占多数,这依赖于 GSH 还原酶,高价硫能否在还原氛的内环境下安稳存在也会是个问题。总之,如何控制酶和底物的互相选择(正交性)以及确保该路径长时间不会出错,可能涉及对整个代码的底层重构。太难了,pass。 磷呢?能量货币 ATP、核酸、磷脂双分子层、激酶等等,和硫一样的问题,涉及底层代码重构,pass。 Cl,离子通道,胃酸的阴离子对,体内大量存在,不适合开发独一无二的信号系统。 最后,只剩 Br,I 和 Se 了。 Br,I 和 Cl,F 一样是卤素,自然界中都不存在单质。C-Cl 键键能 328 kJ/mol,C-Br 键键能 276 kJ/mol,C-I 键键能 240 kJ/mol。键能越低越好操作,需要的能量壁垒低,上下卤素都容易,自然选择项目组的宗旨是能省力做的事情绝不费劲,能跑得动的代码绝不修改,而且 Br 的代码在海洋生命的大模型里跑过(Cell, 2014, Vol. 157, Issue 6, 1380),碘再改改应该就行,那就它吧! 另外,你还需要设计机体获取元素的方式,离子泵的原则是大了小了都不能进,正正好好才可以。碘离子半径大,倘若用溴离子可能过滤不掉一些竞争对手,导致一些奇怪的或者其他地方有用的东西进来而丧失离子通道的特异性。而你设计的碘泵,碘离子的竞争对手是高锝酸盐(TcO4-)、高氯酸盐(ClO4-)、硫氰酸盐(SCN-)这路生理环境中不常见的离子。 但碘和 sp3 碳相连时容易被取代或者消除,多半没等传递到靶器官时就被体内各种亲核物质取代,那就和 sp2 碳的芳环相连吧! 因为碘的强离去能力,Csp3-I 碘代物优先发生的反应 芳环的原材料最好是体内本就充足的,这样不用添加额外代码。氨基酸就不错,有几个带芳环的,那苯丙氨酸,酪氨酸,色氨酸和组氨酸怎么选呢? 连着卤素的 sp2 碳相邻若是氮原子(X-Csp2-N),局部看是个卤代烯胺,反应活性太高,而亲电的物质对机体来说一般都是毒物,会被细胞内的大量存在的 GSH 立即中和清除,所以组氨酸和色氨酸吲哚的吡咯部分 pass。 X=卤素 吲哚的苯环部分和苯丙氨酸的苯环,上碘涉及 C-H 活化和亲电取代,反应条件苛刻,若设计出了活性如此逆天的酶,难保它不会在其他地方胡作非为乱上东西。 这么看酪氨酸的苯酚单元简直眉清目秀,酚羟基邻对位活化苯环利于芳香亲电取代反应(SEAr),电负性高的 N,O,F 还经常有些邻位诱导效应,因此不用太苛刻的活性碘中间体就能轻松实现 Csp2-I 的构建,就它吧。 机体从自然界提取富集的碘是碘离子,碘离子并不像碘单质,能直接和富电子芳环发生 SEAr 反应。但这根本难不倒你,调用广泛存在的过氧化物酶函数,改一改安在新模块里(甲状腺过氧化物酶,thyroperoxidase),专门负责把收集来的碘离子弄成活性碘中间体就成。 载体和反应都选定了,碘的原子半径又大,整个信号分子能变得很“软”,对应的蛋白受体设计起来就非常容易了——增加软软的电子的相互作用就行(受体空腔内部含多个硫原子,形成硫 - 碘次级作用)。不过为了保险起见,还是在分子上多放几个碘,增加容错率,防止随便摄入的一个含碘芳环身体就激动得不要不要的。 3GWS,Crystal Structure of T3-Bound Thyroid Hormone Receptor,受体空腔内部含多个硫原子,形成硫属元素和卤素的相互作用(软亲软,软硬酸碱理论)裹着绿球的紫球是碘原子,黄色折角是硫原子 对了!还要防止与这个模块挨得近的器官把信号分子近水楼台全独吞了,导致近一点儿的器官过度激动,远一些的激动不了。那就设计冗余和局部掉碘的机制打个时间差,以平衡所有器官的反应时间。设计 3 个碘的 T3 活性高、起效快,但持续时间短,掉一个碘就没活性;四个碘的 T4 的活性低、起效慢,但持续时间长,因为还可以脱一个碘变 T3。 如前所述,还需要设计一个对碘代物独一无二的代谢通路,它最好也是稀有的,用到微量元素的,这样不容易出系统级 bug,满足正交性。你看向剩下的 Se 元素,它因为一定的金属性和与 S 元素的相似性一开始并未被你选作信号分子的原材料,但作为掉碘的工具是个不二的选择。 之前项目组为了补充一套可以还原双硫键的 GSH 还原酶的同工酶,开发了含微量元素 Se 的硫氧还蛋白还原酶,TrxR。为此,你做了个SECIS 补丁,此补丁会使核糖体转译时不将 UGA 当作终止密码子,而将其作为编码特殊氨基酸硒半胱氨酸的密码子,最终编码硒蛋白。 于是你用同样的补丁,做了含硒半胱氨酸的脱碘酶(Iodothyronine Deiodinases),用到的化学原理依旧是硫属元素和卤素的相互作用。脱碘酶在组织水平上允许信号分子在给定的细胞内和精确的时空窗口,定时定点地微调,而不会干扰信号分子的血清浓度,目的正是防止上述的器官间的激动不均衡。 DOI: 10.1021/ja210478k ,脱碘酶的一种化学模拟 至此,你通过工作量极小的改造完成了对屎山代码的正交补充,后面就是考虑把这个模块接入 CPU,加些反馈调节机制。这也不是第一次干了,下丘脑 - 垂体 - 肾上腺轴、下丘脑 - 垂体 - 卵巢轴等等都出自项目组的手笔,这个新模块功能强大,理应放置在与 CPU 挨得近的地方,方便调控和管理,那就安在颈部吧(下丘脑 - 垂体 - 甲状腺轴)。 总算能交差了~你为自己精妙绝伦的设计暗爽,可你的老板并不 care,能跑起来就行,祂说。 不过——根据「到目前为止,甲状腺激素是唯一一类含碘的生理物质」的结论和第一性原理的推演,如今遍地的智慧造物们仍能一窥自然选择的魅力。 查看知乎讨论


你是《自然选择》项目组中的一员。

你接到上级指派的新需求:给脊椎动物的屎山代码里加个新系统,要能够控制机体消耗能量、制造蛋白的速度、还要能调节现有的各个系统对各种激素的敏感性,对身体、智力的发育,代谢都要有重要影响。
最高级别响应!!
你看着这闹着玩似的需求陷入沉默,这牵一发动全身的狗屎代码微调都常常跑不动,现在竟要加个 overrule 的管理员级别应用??程序员的命也是命啊。但牛马精神还是驱使你点开了素材库,开始思索怎么用相对简单的方式把以上所有功能加于一个模块上。
既然这个模块的权限那么高,就先把它设定为最大的内分泌腺吧,你思索到。
大致方向有了,那具体的信号传递方式呢?
细胞上的受体容易加,但传递信号的分子最好单一。
因为假如分子多样,那受体也要跟着多样,产生信号分子的器官的复杂性就会提升,信号传递过程中的鲁棒性降低,容易受到屎山系统的干扰而故障。
另外,代谢这个信号分子的通路也最好单独设计,不能是体内广泛存在的酶,不然出 bug 是迟早的事。
但是常规的由 C,H,O,N 组成的分子也容易出 bug。
项目组曾经的战绩是用简单的链烯烃通过一步串联反应产生四五根 C-C 键,构建出了高度刚性、立体专一的甾体和三萜化合物。这神来一笔瞬间提升了分子的复杂度和稳定性,和其他稀稀拉拉、扭来扭去的分子拉开身位。此类骨架用作信号分子(激素),后期只需做些许微调(衍生化),就能满足差异化和不同需求。如此一来,对应的蛋白受体设计起来也省心不少,不需要大幅修改。
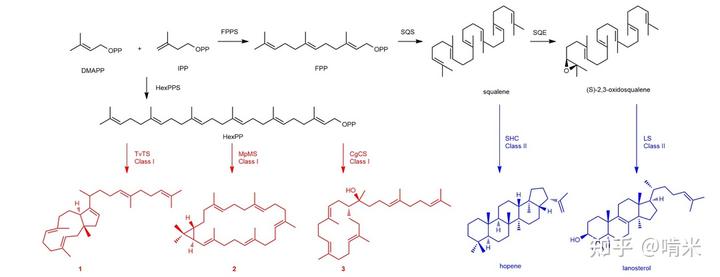
尽管如此,甾体前期的物料准备和后期的衍生化还需要十几二十多步的转化,这还没算上代谢清除过程,简直费时费力。对于调控代谢水平这种即时性很强的需求不能充分的满足。
更要命的是,假如为新模块设计的这个信号分子难以被代谢清除而积蓄,或者受体和其他长得差不多的分子结合,那用户端代谢水平就会不断增加,长此以往早衰 go die。
反之,如果这个由 C,H,O,N 组成的信号分子被广泛存在的氧化还原酶 / 水解酶等等玩意儿截胡,被迫参与了一些针对 O,N 基团的反应,那可能还没来得及传递信号就丧失了功能,用户端代谢水平于是下降,免疫调动不起来,长此以往也 go die。
因此,你开始考虑添加一套完全不同于现有生化反应的新的、能快速反馈和调节的化学反应,正交(互不干扰)的化学不容易出 bug,所以最好加个新的元素,围绕这个元素产生的反应必然和现有体系正交,配得上这个模块管理员级别的地位。
于是在素材库里,你企图为 C,H,O,N 做骨架的小分子增添一些新的化学元素。
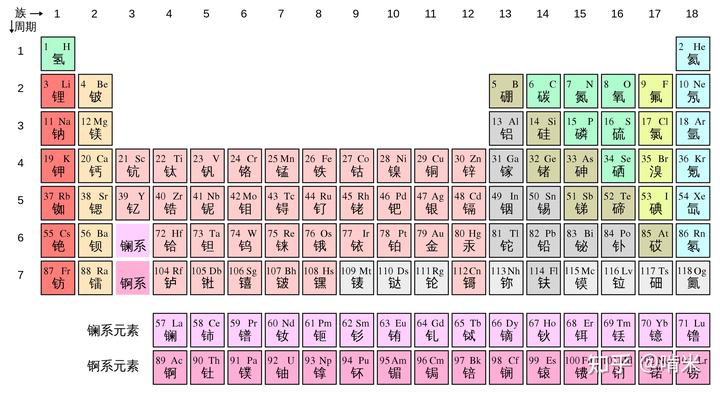
你瞅了眼碱金属(Li, Na, K...)和碱土金属(Ca, Mg..),之前已经被设计组用作离子通道了,pass。
过渡金属(Fe, Zn, Mo, Cu..)多被设计在蛋白里,做信号小分子很难,还有代谢和积蓄的问题,pass。
类金属(B, Si, Ge, As...)容易被氧化还原,而且碳与类金属的化学键很难在复杂的生理环境中稳定。
稀有气体(He, Ne, Ar... )化学惰性,第六七周期的元素基本都是金属,难获取还有放射性更不用考虑。
所以你将目光聚集在非金属上,除了 C,H,O,N 外,还剩 F,S,P,Cl,Se,Br 和 I,碳基生物能和这些元素很好的兼容,正因为能形成稳定的碳 - 杂原子键。
C-F 键能强,难代谢,更难在体内合成,游离的氟源(F2)在自然界中不存在,即便摄入氟离子,也优先和体内的钙离子结合沉积,pass。
S 已经被开发在启动子甲硫氨酸和双硫键里,也就是可以形成蛋白高级构象的半胱氨酸里,还有高能的硫酯键,用在了诸如补体系统的 C3b 里。
如果代谢这个信号分子涉及改变硫元素的氧化态,势必需要氧化还原酶,而这个酶未必不能作用在半胱氨酸上。此外,生理条件下还原型谷胱甘肽(GSH)占多数,这依赖于 GSH 还原酶,高价硫能否在还原氛的内环境下安稳存在也会是个问题。总之,如何控制酶和底物的互相选择(正交性)以及确保该路径长时间不会出错,可能涉及对整个代码的底层重构。太难了,pass。
磷呢?能量货币 ATP、核酸、磷脂双分子层、激酶等等,和硫一样的问题,涉及底层代码重构,pass。
Cl,离子通道,胃酸的阴离子对,体内大量存在,不适合开发独一无二的信号系统。
最后,只剩 Br,I 和 Se 了。
Br,I 和 Cl,F 一样是卤素,自然界中都不存在单质。C-Cl 键键能 328 kJ/mol,C-Br 键键能 276 kJ/mol,C-I 键键能 240 kJ/mol。键能越低越好操作,需要的能量壁垒低,上下卤素都容易,自然选择项目组的宗旨是能省力做的事情绝不费劲,能跑得动的代码绝不修改,而且 Br 的代码在海洋生命的大模型里跑过(Cell, 2014, Vol. 157, Issue 6, 1380),碘再改改应该就行,那就它吧!
另外,你还需要设计机体获取元素的方式,离子泵的原则是大了小了都不能进,正正好好才可以。碘离子半径大,倘若用溴离子可能过滤不掉一些竞争对手,导致一些奇怪的或者其他地方有用的东西进来而丧失离子通道的特异性。而你设计的碘泵,碘离子的竞争对手是高锝酸盐(TcO4-)、高氯酸盐(ClO4-)、硫氰酸盐(SCN-)这路生理环境中不常见的离子。
但碘和 sp3 碳相连时容易被取代或者消除,多半没等传递到靶器官时就被体内各种亲核物质取代,那就和 sp2 碳的芳环相连吧!
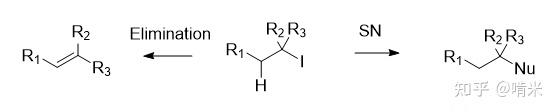
芳环的原材料最好是体内本就充足的,这样不用添加额外代码。氨基酸就不错,有几个带芳环的,那苯丙氨酸,酪氨酸,色氨酸和组氨酸怎么选呢?
连着卤素的 sp2 碳相邻若是氮原子(X-Csp2-N),局部看是个卤代烯胺,反应活性太高,而亲电的物质对机体来说一般都是毒物,会被细胞内的大量存在的 GSH 立即中和清除,所以组氨酸和色氨酸吲哚的吡咯部分 pass。
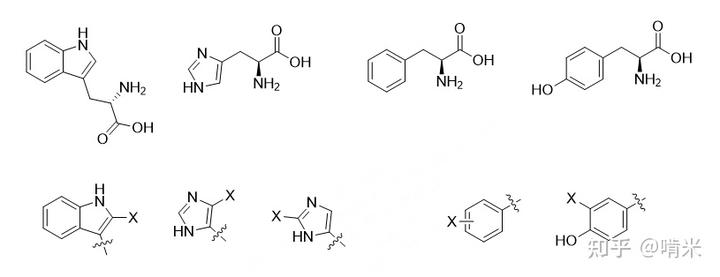
吲哚的苯环部分和苯丙氨酸的苯环,上碘涉及 C-H 活化和亲电取代,反应条件苛刻,若设计出了活性如此逆天的酶,难保它不会在其他地方胡作非为乱上东西。
这么看酪氨酸的苯酚单元简直眉清目秀,酚羟基邻对位活化苯环利于芳香亲电取代反应(SEAr),电负性高的 N,O,F 还经常有些邻位诱导效应,因此不用太苛刻的活性碘中间体就能轻松实现 Csp2-I 的构建,就它吧。
机体从自然界提取富集的碘是碘离子,碘离子并不像碘单质,能直接和富电子芳环发生 SEAr 反应。但这根本难不倒你,调用广泛存在的过氧化物酶函数,改一改安在新模块里(甲状腺过氧化物酶,thyroperoxidase),专门负责把收集来的碘离子弄成活性碘中间体就成。
载体和反应都选定了,碘的原子半径又大,整个信号分子能变得很“软”,对应的蛋白受体设计起来就非常容易了——增加软软的电子的相互作用就行(受体空腔内部含多个硫原子,形成硫 - 碘次级作用)。不过为了保险起见,还是在分子上多放几个碘,增加容错率,防止随便摄入的一个含碘芳环身体就激动得不要不要的。
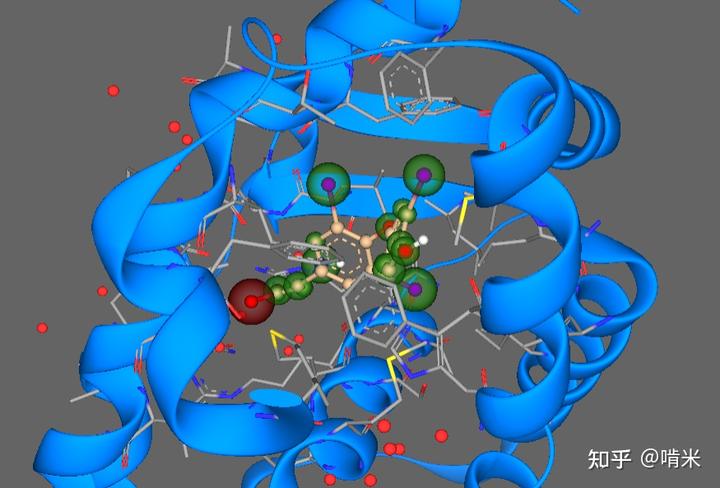
对了!还要防止与这个模块挨得近的器官把信号分子近水楼台全独吞了,导致近一点儿的器官过度激动,远一些的激动不了。那就设计冗余和局部掉碘的机制打个时间差,以平衡所有器官的反应时间。设计 3 个碘的 T3 活性高、起效快,但持续时间短,掉一个碘就没活性;四个碘的 T4 的活性低、起效慢,但持续时间长,因为还可以脱一个碘变 T3。
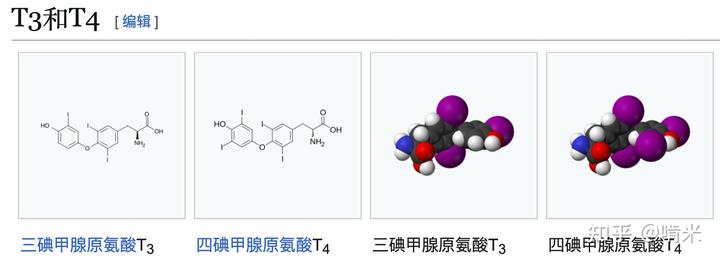
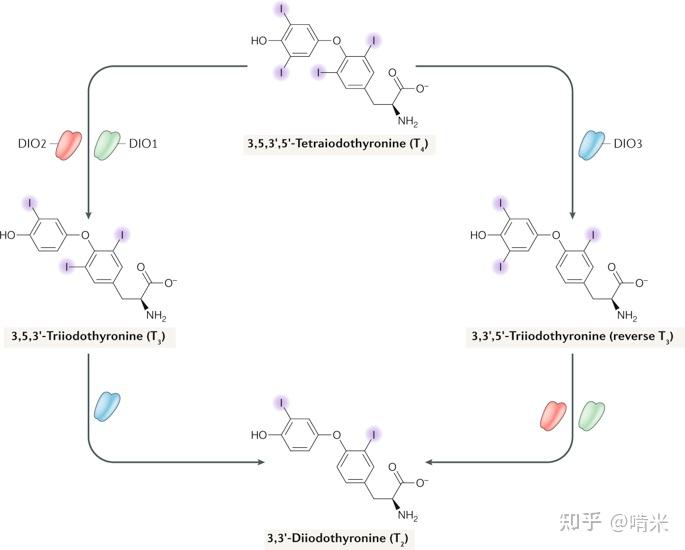
如前所述,还需要设计一个对碘代物独一无二的代谢通路,它最好也是稀有的,用到微量元素的,这样不容易出系统级 bug,满足正交性。你看向剩下的 Se 元素,它因为一定的金属性和与 S 元素的相似性一开始并未被你选作信号分子的原材料,但作为掉碘的工具是个不二的选择。
之前项目组为了补充一套可以还原双硫键的 GSH 还原酶的同工酶,开发了含微量元素 Se 的硫氧还蛋白还原酶,TrxR。为此,你做了个SECIS 补丁,此补丁会使核糖体转译时不将 UGA 当作终止密码子,而将其作为编码特殊氨基酸硒半胱氨酸的密码子,最终编码硒蛋白。
于是你用同样的补丁,做了含硒半胱氨酸的脱碘酶(Iodothyronine Deiodinases),用到的化学原理依旧是硫属元素和卤素的相互作用。脱碘酶在组织水平上允许信号分子在给定的细胞内和精确的时空窗口,定时定点地微调,而不会干扰信号分子的血清浓度,目的正是防止上述的器官间的激动不均衡。
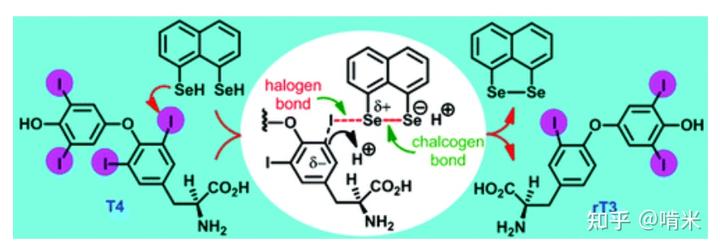
至此,你通过工作量极小的改造完成了对屎山代码的正交补充,后面就是考虑把这个模块接入 CPU,加些反馈调节机制。这也不是第一次干了,下丘脑 - 垂体 - 肾上腺轴、下丘脑 - 垂体 - 卵巢轴等等都出自项目组的手笔,这个新模块功能强大,理应放置在与 CPU 挨得近的地方,方便调控和管理,那就安在颈部吧(下丘脑 - 垂体 - 甲状腺轴)。
总算能交差了~你为自己精妙绝伦的设计暗爽,可你的老板并不 care,能跑起来就行,祂说。
不过——根据「到目前为止,甲状腺激素是唯一一类含碘的生理物质」的结论和第一性原理的推演,如今遍地的智慧造物们仍能一窥自然选择的魅力。












