



























【地球演义】偏师劲旅(中):死海魅影
书接前文,上回井蛙留了个扣子,说副爬行动物中有两个“明星”类群:中龙类(Mesosauria)和巨齿龙类(Pareiasauria)。这回我们先介绍中龙。 中龙类生活在二叠纪刚刚开始不久(大约 2.99 亿到 2.75 亿年前)。在巴西,乌拉圭,纳米比亚和南非一共发现了三种中龙类的化石。它们是中龙属的纳米比亚中龙(Mesosaurus tenuidens),方胸龙属的凸方胸龙(Stereosternum tumidum),和巴西龙属的圣保罗巴西龙(Brazilosaurus sanpauloensis)。 中龙类三个属代表物种复原图。上:纳米比亚中龙;中:凸方胸龙;下:圣保罗巴西龙。可以看到它们在体型,身体比例和骨骼结构方面有着细微的差别。图片来源自网络。 一眼就可以看出,中龙类是高度水栖的类群:柔长的躯体,细弱的四肢,超过体长一半的大尾巴,灵活的脖颈,还有高度特化的口鼻和牙齿。这些特征明显比同时代的其他水栖羊膜类:齐椎蜥(Spinoaequalis),坦噶蜥(Tamgasaurus),霍瓦蜥(Hovasaurus)和克劳迪欧蜥(Claudiosaurus,参见第二百五回 水陆空 V1.0)更加偏向水生。感觉上,中龙类有点“超前”,即使把它们放到中生代的海爬中也没什么违和感。 纳米比亚中龙骨骼复原图(年轻的成体)。中龙类的体型不算太大,成体体长一米左右,最大的也不到到 2 米。图片来源自[1]。 凸方胸龙骨骼化石。中龙类的体态很像中生代的一些海爬,比如化石市场上到处可见的贵州龙(Keichousaurus,三叠纪的回目再详细介绍),但生存年代却要至少早上 9000 万年。图片来源见水印。 圣保罗巴西龙(发现地:Montividiu)化石。和另外两种中龙相比,它的身体更苗条,头部更小,颈部也更长。图片来源自[2]。 目前研究最深入的是纳米比亚中龙。这种动物留下了极其丰富的骨骼标本,包括软体组织的印痕和不同发育阶段的幼体。 这里插一句,乌拉圭的 Mangrullo 化石群是发现中龙化石的宝地,这里出土了许多保存程度超乎想象的标本。比如上图中的纳米比亚中龙下颌化石,连贯穿在骨骼和牙齿中的神经(黑色箭头标示)和血管(灰色箭头标示)都清晰可辨。图片来源自[3]。 但即使如此,人们仍然被一些最基本的问题困惑着。比如中龙类的分类地位。可以确定中龙类起源于某种陆栖的羊膜动物。演化出如此明显的水栖特征肯定不是一朝一夕的事。但遗憾的是,目前还没有发现关于中龙祖先和过渡类型的任何蛛丝马迹。而这样特化的动物,只靠形态学进行分类又是不大靠得住的。一直以来,人们把中龙类看做是副爬行动物中一个非常古老,而且早早分化出去的旁支(参见上回的副爬行动物谱系图)。但一些新的研究显示,中龙类的祖先也许可以追溯到副爬行动物和真爬行动物分家之前。 一种关于早期羊膜动物谱系发育的理论(图片来源自[4]),中龙被放在蜥形纲最基干的位置。但这个理论认为龟鳖类属于副爬行动物,甚至把副爬行动物都放到“双孔类”的演化支下,总之挺颠覆的,接受度不高。本文仍然采用中龙类属于副爬行动物的传统分类。 从身体结构上看,中龙类在陆地上运动已经很吃力了。它一生的大多数时间(如果不是终生水栖的话)都应该是在水中渡过。 各个发育阶段的纳米比亚中龙后足骨骼结构。它的趾骨细长,五趾分开,趾间应该长有蹼辅助划水。图片来源自[5]。 纳米比亚中龙成体后肢骨骼结构。它的腿骨和骨盆非常细弱,在陆地上无法支撑身体。大概最多也就是像海龟一样费力地匍匐爬行吧。图片来源自[5],标尺长度 1 厘米。 中龙化石给人印象最深刻的是那张尖细的长嘴,上下颚长满锋利的牙齿,这些长牙伸出口外,交叉错叠,组成一张看起来十分凶残的大杀器。 纳米比亚中龙头骨化石和复原图。中龙类的眼睛很大,视力应该不错。头骨下后侧也有一对小小的颞颥孔。这样的牙齿在二叠纪四足动物中绝无仅有。图片来源自[6],标尺长度 1 厘米。 相信很多朋友已经在脑海中描绘这样的图景:几条中龙在水中轻轻扭摆,懒洋洋地游荡在鱼群四周。看准时机后,大尾巴猛地发力,修长的身体就像鱼雷一样弹射出去。大嘴一张咬住一条大鱼,锋利的牙齿牢牢钉进猎物的身体,长颈一缩一探,便吞了下去。跟着身子轻转,目光又回到惊逃的鱼群,开始物色下一个猎物…… 是的,井蛙刚看到中龙化石的时候也是这么想的。但是后来才知道,这个画面根本不会出现。原因很简单,这是一张中龙生活环境的复原图(图片来源自网络): 你可以看到中龙在水中遨游,还趴在岸上晒太阳(也许会吧),水边生长着茂密的植被。但是注意:这片水里——根本——没有鱼!种种证据显示,中龙确实是这片水域的霸主,然而它们统治的水体,却是异乎寻常的贫瘠!(注意右下那个圆形的动物,等下说它是什么)。 研究者在乌拉圭 Mangrullo 早二叠世化石群找到了上百具中龙化石。上图标示了这里出土的化石种类和出现地层。对比左上的图例,可以看到在发现大量中龙骨骼的地层中,有植物叶片和昆虫翅膀化石,这明显是来自岸边的陆地植被。但是却没有发现珊瑚,海绵和苔藓虫,也没有腕足动物,没有腹足类和双壳类,没有菊石,没有鱼类,也没有离片椎类,壳椎类和其他羊膜动物。这是怎样一片空旷死寂的水体啊。图片来源自[3]。 唯一和中龙大量伴生的,是虾,大量的虾,就是在前面的回目(参见第二百二十二回 虾啊,真虾,真是虾,是真虾……攀缘的井蛙:【地球演义】虾啊,真虾,真是虾,是真虾……)介绍过的,二叠纪最繁盛的甲壳类——臀头虾(Pygocephalomorpha)。上面复原图里那个圆溜溜的动物就是臀头虾。 Mangrullo 化石群生态系统中动物类型统计表。这里发现了 58 条纳米比亚中龙的骨骼,57 条凸方胸龙的骨骼。还有三种臀头虾:A 种发现了 156 个个体,是绝对的优势种类;B 种发现了 19 个个体,C 种发现了 1 个个体。然后,就没有了,什么也没有了。图表来源自[3]。 在我们的认知里,无论海洋还是淡水河湖,都不会有这么奇怪的动物组成。那么中龙类到底生活在什么样的环境里呢?一种比较圆满的解释是,中龙类很可能生活在内陆的盐湖(Hypersaline sea)中。盐湖是内流河(Continental river)的终点,河水不断注入,带来溶解在其中的矿物质。随着湖水不断蒸发,不断浓缩,最终达到很高的盐度,令绝大多数生物都无法在其中生存——就像现在的死海那样。 中龙化石发现地(红点)在冈瓦纳古陆的对应位置。在二叠纪早期,非洲(1)和南美(2)是连在一起的,中间的海峡并不存在。所以中龙类并不是生活在海边,而是内陆。图片来源自网络。 现代盐湖中也往往存在非常单一的生态系统。只有一种或寥寥几种藻类能在高盐度的湖水中生存,耐盐的小型无脊椎动物,比如卤虫(Brine shrimp,上图)和盐水蝇(Brine fly,下图)组成了食物链的下一个环节。因为没有其他动物与之竞争,它们的数量往往异常庞大。 Mangrullo 化石群发现的臀头虾类化石。它们很可能在二叠纪的盐湖中占据着今天卤虫或者盐水蝇的生态位,是某种远古耐盐藻类唯一的消费者。图片来源自[3]。 从化石证据来看,这些臀头虾的小日子过得还挺滋润,因为没有鱼类去捕食它们。上图是 Mangrullo 化石群发现的正在交配的臀头虾化石。它们就在这一汪齁死人的咸汤里快乐地生息繁衍。图片来源自[3]。 可以推测,机缘巧合之下,中龙类的陆生祖先发现这片盐湖。湖里没有鲨鱼也没有凶猛的离片椎类,有的是吃不完的虾,虽然个头小了一点,但数量绝对管够。经过一代代的选择与淘汰,在解决了水盐代谢的难题(不知道中龙会不会像海龟和咸水鳄一样淌眼泪)之后,它们的后代成为这片高盐水域的顶级掠食者。 现代盐湖里的小型无脊椎动物也会吸引特定的脊椎动物前来取食。比如上图中的火烈鸟,就用特殊的鸟喙滤取湖水中微小的卤虫。很可能在 2.9 亿年前的盐湖中,甲壳类和蜥形纲的前辈们就已经建立起类似的食物链了。图片来源自网络。 有许多中龙化石保留了腹部的内容物,从中可以清晰地分辨出臀头虾的残骸。如此看来,中龙略显恐怖的长牙其实主要用来捞取和咬碎这些小型甲壳动物。 一条保存非常完好的纳米比亚中龙前半身化石(A 图)。在它的肩胛骨下方有一个完整的大臀头虾(B 图箭头标示),那是它吞下的最后一口食物,可能是在掩埋的过程中从食道里挤出来的。图片来源自[1]。 除了臀头虾,研究者还在中龙的腹腔,粪便和呕吐物里发现了其他动物残骸——同类的幼体。 Mangrullo 化石群发现的纳米比亚中龙化石,在腹腔中发现了食物残留的印痕(A)。放大图见 B 和 C,其中箭头标示的是臀头虾的外壳,锐角三角形指示的是中龙幼体的碎骨头。图片来源自[3] 。 一些在 Mangrullo 化石群发现的纳米比亚中龙粪化石(Coprolites)。A 图和 C 图中只有臀头虾的外骨骼;B 图中则有非常小的中龙幼体骨骼碎片(红线标示);D 图中不但有骨骼碎片,而且还有一枚牙齿(红色箭头标示)。图片来源自[1],标尺长度 1 厘米。 一些 Mangrullo 化石群发现的呕吐物化石(Regurgitalites),可以分辨出其中混杂着 中龙幼体的颈椎(cv),股骨(f),肋骨(fr),颅骨(fskb),牙齿(te)和其他骨骼(ub)的碎片。因为这里没有发现其他大型脊椎动物,所以可以基本确定来自中龙。看来它们会像许多现代动物一样,把不能消化,也不太好从肠道排除的硬质杂物呕吐出来。图片来源自[1],标尺长度 1 厘米。 另一处中龙化石发现点:巴西 Iratí化石群(位于大名鼎鼎的巴拉纳盆地)发现的中龙粪化石和肠内容物化石(Cololites)。A 图是一个粪堆。B 图粪里有一枚非常小的牙齿(标尺长度 1 厘米,放大后如 C 图,标尺长度 5 毫米)。D 图粪里有许多骨头碎片(箭头标示)。E 图是粪化石截面的 SEM 照片,可以看到许多臀头虾壳和中龙幼体的碎骨纠缠挤压在一起,这是中龙的两种主要食物。图片来源自[1]。 有人认为这是中龙同类相食的证据,也有人认为中龙会啃食同类的尸体。井蛙更倾向于前者。这么多的化石证据表明吃掉幼体的情况非常普遍,很难用食腐来解释。很可能因为 2 厘米的臀头虾对 1 米多长的中龙成体来说,还是太小了。中龙只能靠自己来填补食物链中间的空缺,在贫瘠的盐湖里建立起这样一条能量传递途径:藻类→臀头虾→中龙幼体→中龙亚成体→中龙成体。至于死去中龙的尸体,大概臀头虾会吃掉一部分,其他的中龙会吃掉一部分,剩下的残骸就交给微生物分解。腌在这种浓盐水里,尸体的腐烂速度会变得缓慢,就有更大的几率被湖底的沉积物覆盖掩埋。这也解释了为什么会有这么多保存完好的化石出土。 最为惊艳的化石要数中龙的胚胎。这也是人们发现的最早的羊膜动物胚胎化石。 Mangrullo 化石群发现的纳米比亚中龙化石。在成体腹部左侧有一个完整的胚胎骨骼(A 图)。B 图是胚胎和刚孵化幼体的体型对比。图片来源自[7]。 上图中的中龙胚胎骨骼结构和复原图。这个宝宝蜷缩成一团,但化石上没有卵壳的痕迹,它很可能是包裹在薄薄的卵膜里。图片来源自[7],标尺长度 5 毫米。 在这个胚胎的长嘴上发现了一个钉子状的小突起(箭头标示)。有人认为这是它的卵齿(egg tooth),用来顶破卵壳或卵膜的。图片来源自[7],标尺长度 1 厘米。 巴西 Iratí化石群发现的纳米比亚中龙化石。在它的腹腔里也有一个发育成型的胚胎(如 D 图所示)。图片来源自[7]。 另外两块中龙化石。它们的腹腔里也发现了比较完整的幼体骨架(骨骼排列有序,基本可以排除是咬碎后吃掉的)。这些幼体比前面的胚胎大一些,可能更临近出生。图片来源自[7],标尺长度 1 厘米。 这些化石证据表明,中龙类的胚胎会在母体内充分发育,这也是对水栖生活的一种适应。它们要么孵化时间很短,说不定在水中产卵,幼体出生后马上用卵齿划开卵膜,开始自由生活。或者直接产下幼体(就像中生代的鱼龙那样)。如果是这样的话,那中龙确实没有理由再爬上岸去了


书接前文,上回井蛙留了个扣子,说副爬行动物中有两个“明星”类群:中龙类(Mesosauria)和巨齿龙类(Pareiasauria)。这回我们先介绍中龙。
中龙类生活在二叠纪刚刚开始不久(大约 2.99 亿到 2.75 亿年前)。在巴西,乌拉圭,纳米比亚和南非一共发现了三种中龙类的化石。它们是中龙属的纳米比亚中龙(Mesosaurus tenuidens),方胸龙属的凸方胸龙(Stereosternum tumidum),和巴西龙属的圣保罗巴西龙(Brazilosaurus sanpauloensis)。
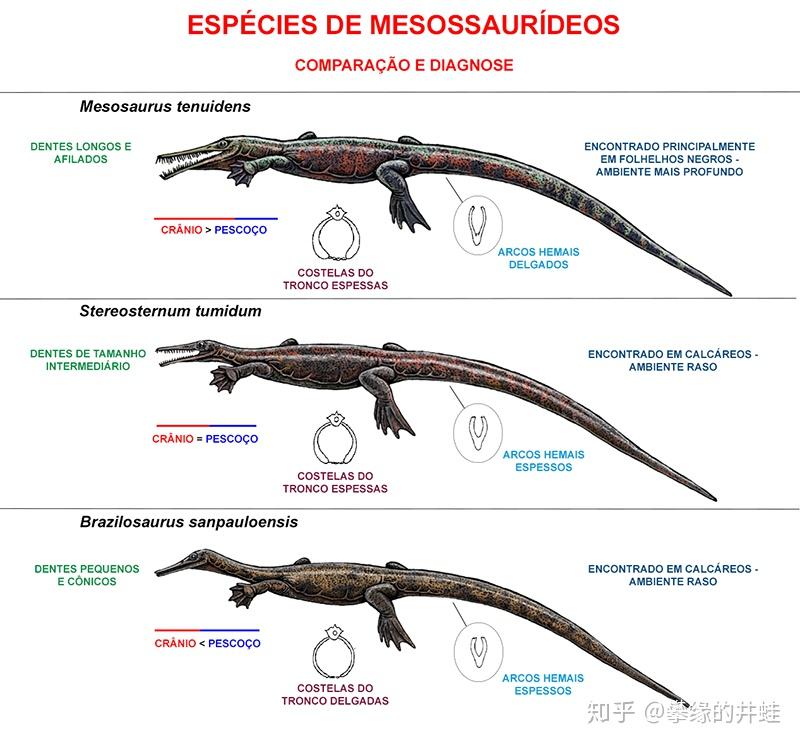
中龙类三个属代表物种复原图。上:纳米比亚中龙;中:凸方胸龙;下:圣保罗巴西龙。可以看到它们在体型,身体比例和骨骼结构方面有着细微的差别。图片来源自网络。
一眼就可以看出,中龙类是高度水栖的类群:柔长的躯体,细弱的四肢,超过体长一半的大尾巴,灵活的脖颈,还有高度特化的口鼻和牙齿。这些特征明显比同时代的其他水栖羊膜类:齐椎蜥(Spinoaequalis),坦噶蜥(Tamgasaurus),霍瓦蜥(Hovasaurus)和克劳迪欧蜥(Claudiosaurus,参见第二百五回 水陆空 V1.0)更加偏向水生。感觉上,中龙类有点“超前”,即使把它们放到中生代的海爬中也没什么违和感。

纳米比亚中龙骨骼复原图(年轻的成体)。中龙类的体型不算太大,成体体长一米左右,最大的也不到到 2 米。图片来源自[1]。

凸方胸龙骨骼化石。中龙类的体态很像中生代的一些海爬,比如化石市场上到处可见的贵州龙(Keichousaurus,三叠纪的回目再详细介绍),但生存年代却要至少早上 9000 万年。图片来源见水印。
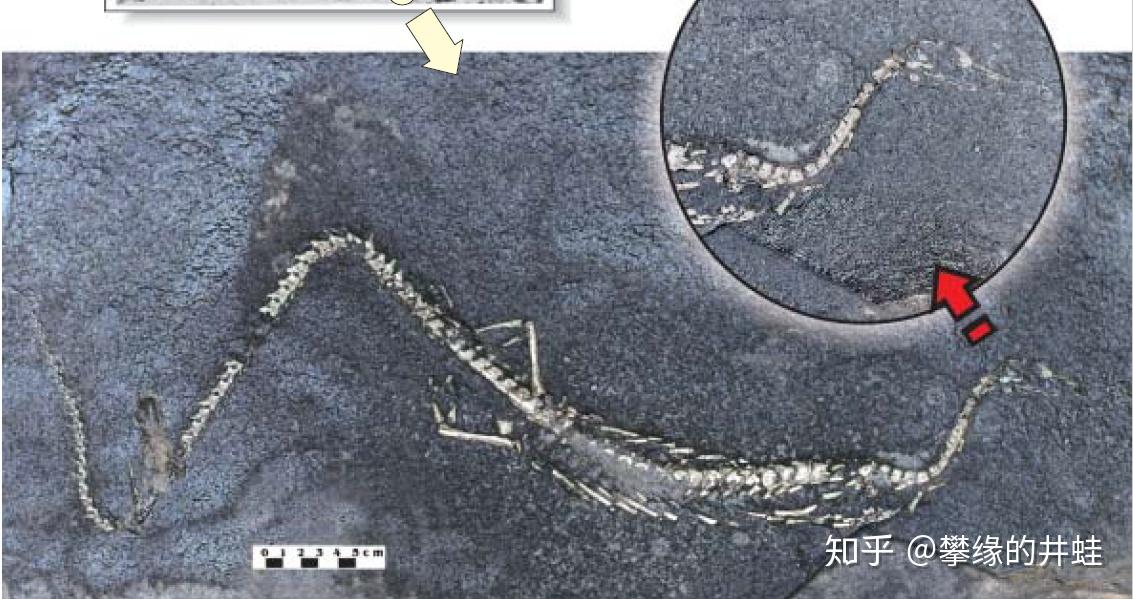
圣保罗巴西龙(发现地:Montividiu)化石。和另外两种中龙相比,它的身体更苗条,头部更小,颈部也更长。图片来源自[2]。
目前研究最深入的是纳米比亚中龙。这种动物留下了极其丰富的骨骼标本,包括软体组织的印痕和不同发育阶段的幼体。
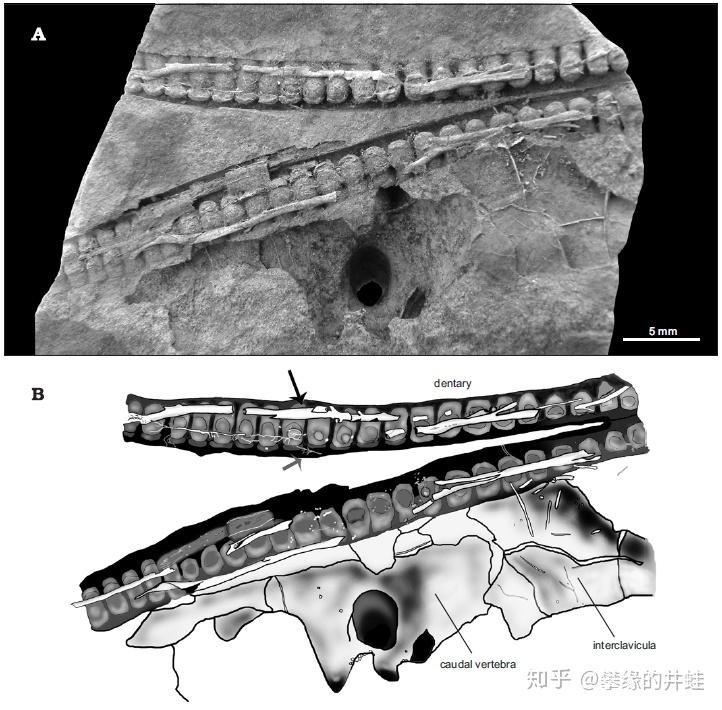
这里插一句,乌拉圭的 Mangrullo 化石群是发现中龙化石的宝地,这里出土了许多保存程度超乎想象的标本。比如上图中的纳米比亚中龙下颌化石,连贯穿在骨骼和牙齿中的神经(黑色箭头标示)和血管(灰色箭头标示)都清晰可辨。图片来源自[3]。
但即使如此,人们仍然被一些最基本的问题困惑着。比如中龙类的分类地位。可以确定中龙类起源于某种陆栖的羊膜动物。演化出如此明显的水栖特征肯定不是一朝一夕的事。但遗憾的是,目前还没有发现关于中龙祖先和过渡类型的任何蛛丝马迹。而这样特化的动物,只靠形态学进行分类又是不大靠得住的。一直以来,人们把中龙类看做是副爬行动物中一个非常古老,而且早早分化出去的旁支(参见上回的副爬行动物谱系图)。但一些新的研究显示,中龙类的祖先也许可以追溯到副爬行动物和真爬行动物分家之前。
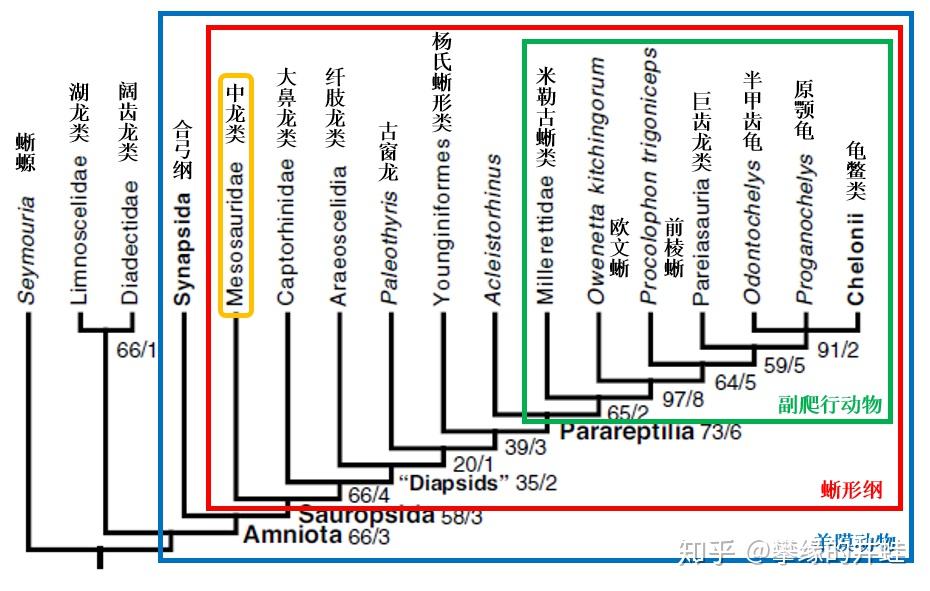
一种关于早期羊膜动物谱系发育的理论(图片来源自[4]),中龙被放在蜥形纲最基干的位置。但这个理论认为龟鳖类属于副爬行动物,甚至把副爬行动物都放到“双孔类”的演化支下,总之挺颠覆的,接受度不高。本文仍然采用中龙类属于副爬行动物的传统分类。
从身体结构上看,中龙类在陆地上运动已经很吃力了。它一生的大多数时间(如果不是终生水栖的话)都应该是在水中渡过。

各个发育阶段的纳米比亚中龙后足骨骼结构。它的趾骨细长,五趾分开,趾间应该长有蹼辅助划水。图片来源自[5]。

纳米比亚中龙成体后肢骨骼结构。它的腿骨和骨盆非常细弱,在陆地上无法支撑身体。大概最多也就是像海龟一样费力地匍匐爬行吧。图片来源自[5],标尺长度 1 厘米。
中龙化石给人印象最深刻的是那张尖细的长嘴,上下颚长满锋利的牙齿,这些长牙伸出口外,交叉错叠,组成一张看起来十分凶残的大杀器。
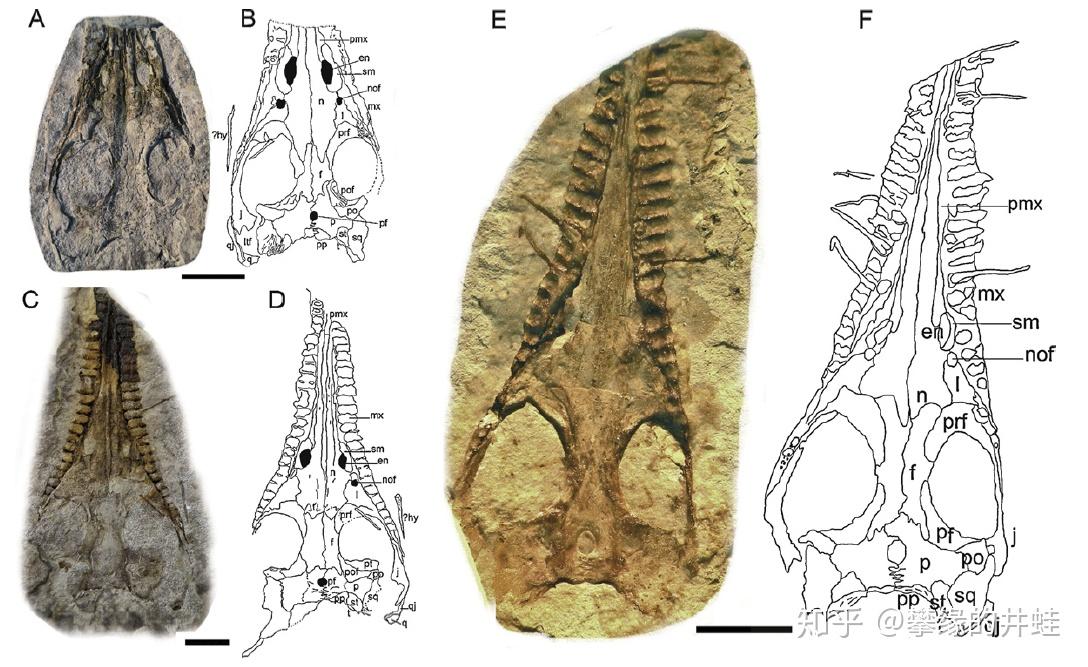
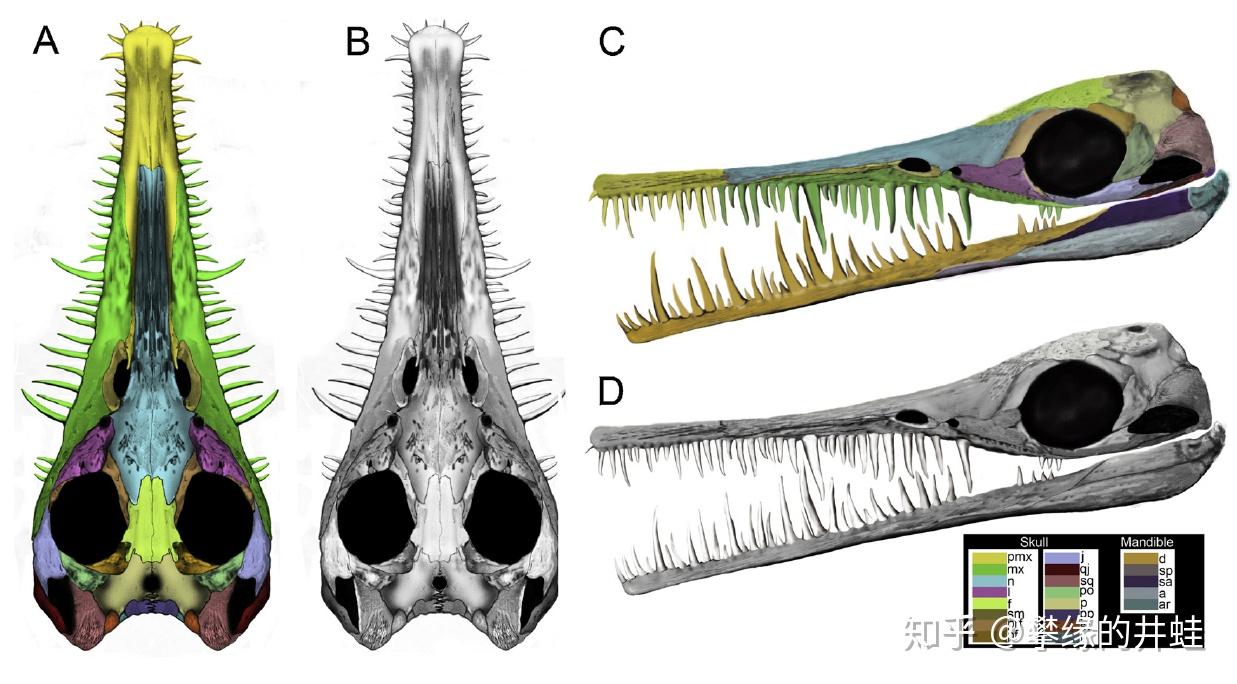
纳米比亚中龙头骨化石和复原图。中龙类的眼睛很大,视力应该不错。头骨下后侧也有一对小小的颞颥孔。这样的牙齿在二叠纪四足动物中绝无仅有。图片来源自[6],标尺长度 1 厘米。
相信很多朋友已经在脑海中描绘这样的图景:几条中龙在水中轻轻扭摆,懒洋洋地游荡在鱼群四周。看准时机后,大尾巴猛地发力,修长的身体就像鱼雷一样弹射出去。大嘴一张咬住一条大鱼,锋利的牙齿牢牢钉进猎物的身体,长颈一缩一探,便吞了下去。跟着身子轻转,目光又回到惊逃的鱼群,开始物色下一个猎物……
是的,井蛙刚看到中龙化石的时候也是这么想的。但是后来才知道,这个画面根本不会出现。原因很简单,这是一张中龙生活环境的复原图(图片来源自网络):

你可以看到中龙在水中遨游,还趴在岸上晒太阳(也许会吧),水边生长着茂密的植被。但是注意:这片水里——根本——没有鱼!种种证据显示,中龙确实是这片水域的霸主,然而它们统治的水体,却是异乎寻常的贫瘠!(注意右下那个圆形的动物,等下说它是什么)。

研究者在乌拉圭 Mangrullo 早二叠世化石群找到了上百具中龙化石。上图标示了这里出土的化石种类和出现地层。对比左上的图例,可以看到在发现大量中龙骨骼的地层中,有植物叶片和昆虫翅膀化石,这明显是来自岸边的陆地植被。但是却没有发现珊瑚,海绵和苔藓虫,也没有腕足动物,没有腹足类和双壳类,没有菊石,没有鱼类,也没有离片椎类,壳椎类和其他羊膜动物。这是怎样一片空旷死寂的水体啊。图片来源自[3]。
唯一和中龙大量伴生的,是虾,大量的虾,就是在前面的回目(参见第二百二十二回 虾啊,真虾,真是虾,是真虾……
攀缘的井蛙:【地球演义】虾啊,真虾,真是虾,是真虾……
)介绍过的,二叠纪最繁盛的甲壳类——臀头虾(Pygocephalomorpha)。上面复原图里那个圆溜溜的动物就是臀头虾。
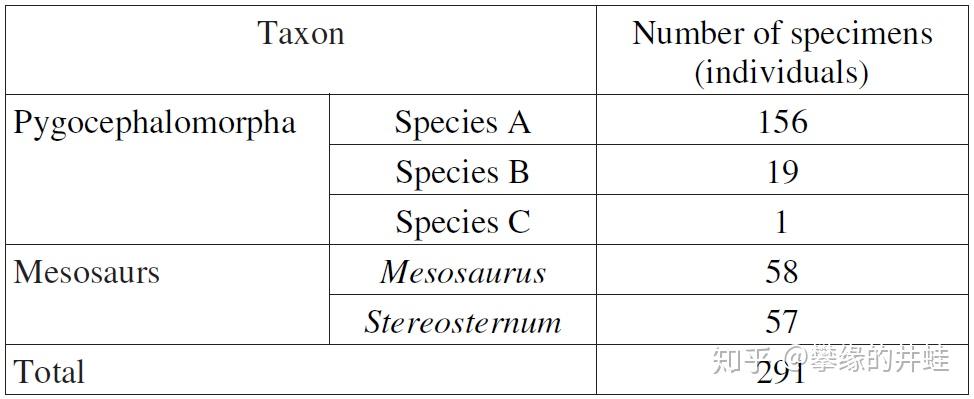
Mangrullo 化石群生态系统中动物类型统计表。这里发现了 58 条纳米比亚中龙的骨骼,57 条凸方胸龙的骨骼。还有三种臀头虾:A 种发现了 156 个个体,是绝对的优势种类;B 种发现了 19 个个体,C 种发现了 1 个个体。然后,就没有了,什么也没有了。图表来源自[3]。
在我们的认知里,无论海洋还是淡水河湖,都不会有这么奇怪的动物组成。那么中龙类到底生活在什么样的环境里呢?一种比较圆满的解释是,中龙类很可能生活在内陆的盐湖(Hypersaline sea)中。盐湖是内流河(Continental river)的终点,河水不断注入,带来溶解在其中的矿物质。随着湖水不断蒸发,不断浓缩,最终达到很高的盐度,令绝大多数生物都无法在其中生存——就像现在的死海那样。
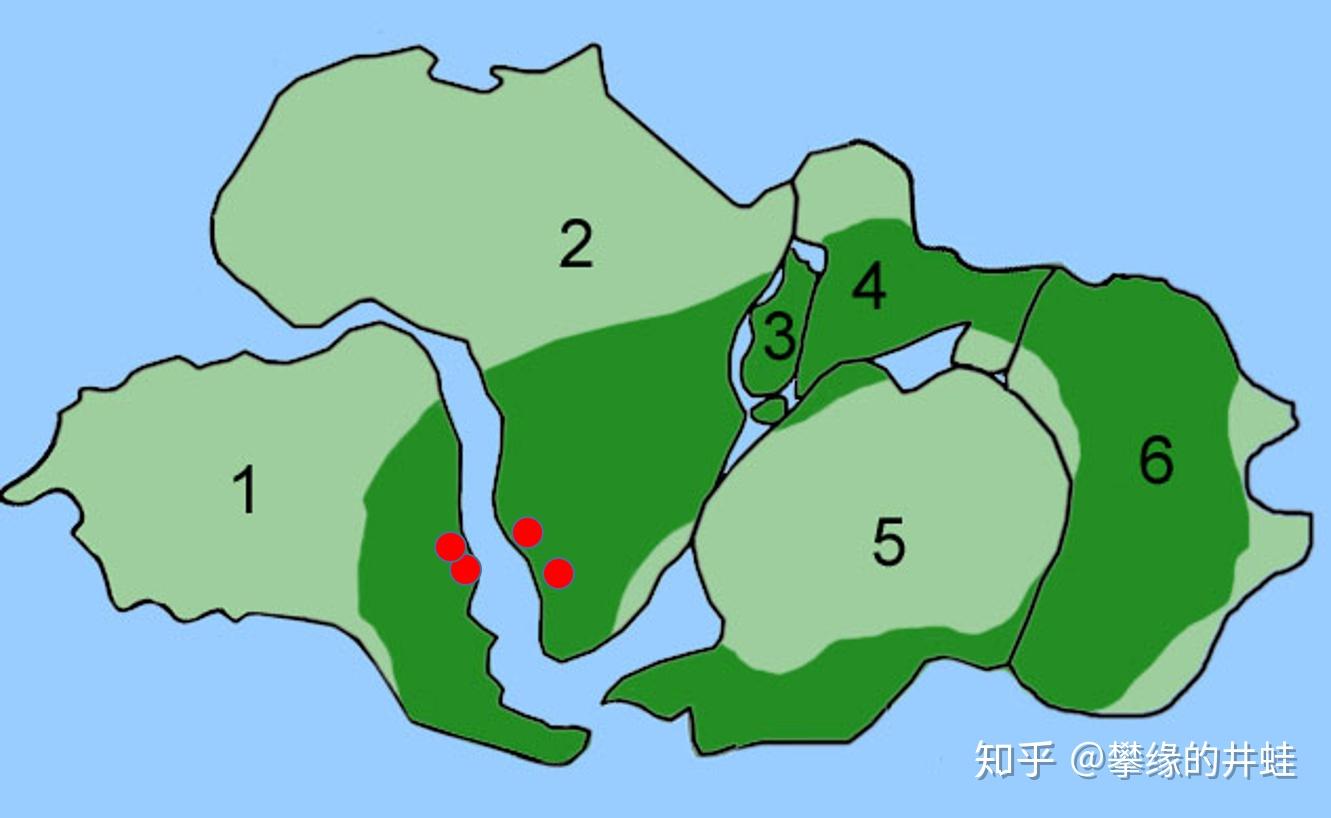
中龙化石发现地(红点)在冈瓦纳古陆的对应位置。在二叠纪早期,非洲(1)和南美(2)是连在一起的,中间的海峡并不存在。所以中龙类并不是生活在海边,而是内陆。图片来源自网络。
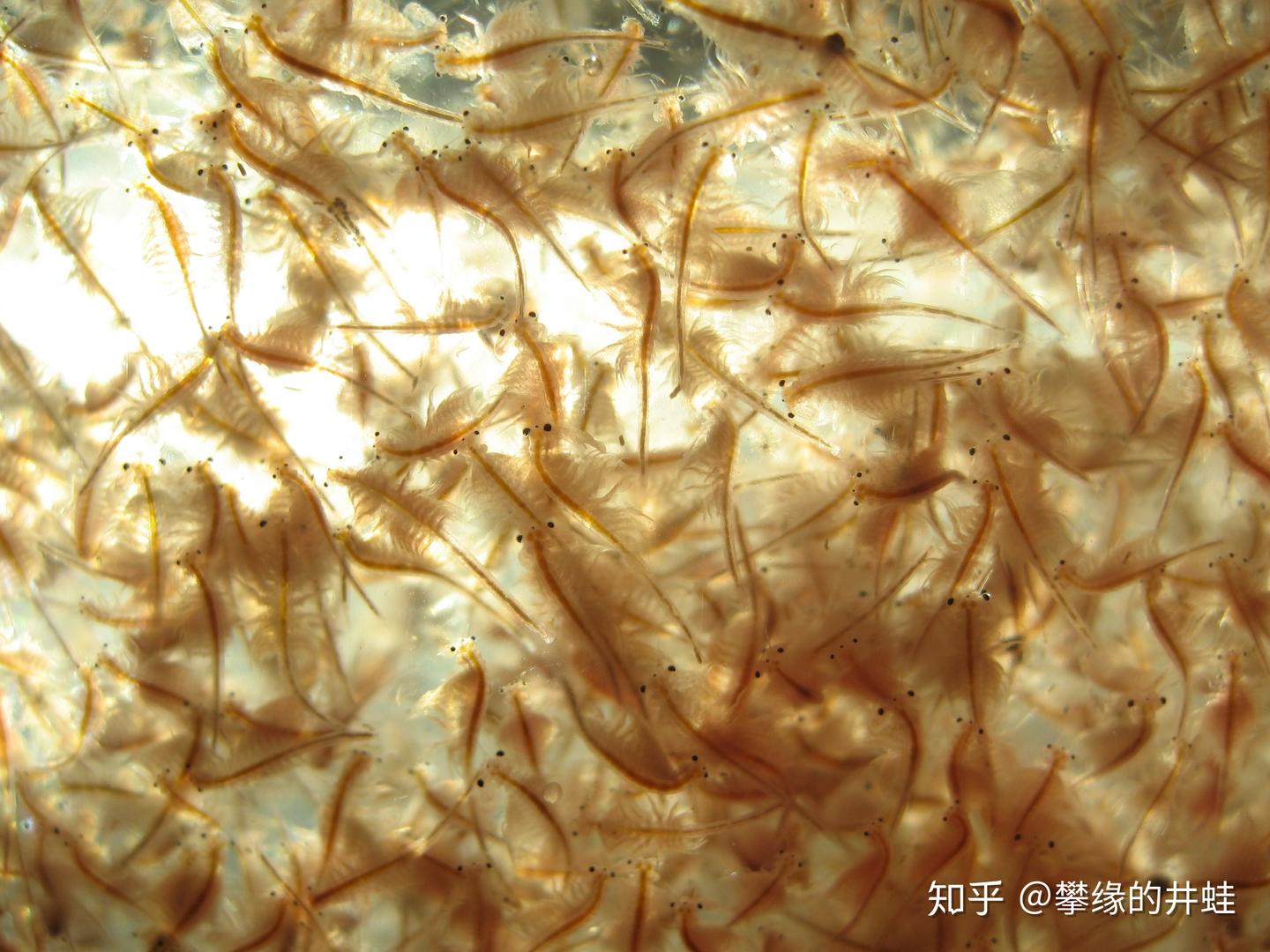

现代盐湖中也往往存在非常单一的生态系统。只有一种或寥寥几种藻类能在高盐度的湖水中生存,耐盐的小型无脊椎动物,比如卤虫(Brine shrimp,上图)和盐水蝇(Brine fly,下图)组成了食物链的下一个环节。因为没有其他动物与之竞争,它们的数量往往异常庞大。
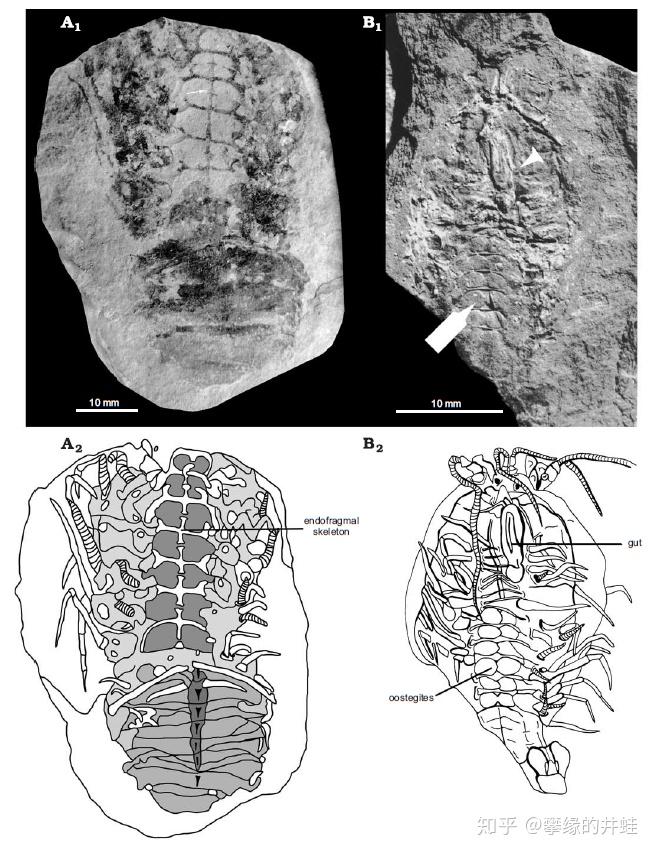
Mangrullo 化石群发现的臀头虾类化石。它们很可能在二叠纪的盐湖中占据着今天卤虫或者盐水蝇的生态位,是某种远古耐盐藻类唯一的消费者。图片来源自[3]。
从化石证据来看,这些臀头虾的小日子过得还挺滋润,因为没有鱼类去捕食它们。上图是 Mangrullo 化石群发现的正在交配的臀头虾化石。它们就在这一汪齁死人的咸汤里快乐地生息繁衍。图片来源自[3]。
可以推测,机缘巧合之下,中龙类的陆生祖先发现这片盐湖。湖里没有鲨鱼也没有凶猛的离片椎类,有的是吃不完的虾,虽然个头小了一点,但数量绝对管够。经过一代代的选择与淘汰,在解决了水盐代谢的难题(不知道中龙会不会像海龟和咸水鳄一样淌眼泪)之后,它们的后代成为这片高盐水域的顶级掠食者。

现代盐湖里的小型无脊椎动物也会吸引特定的脊椎动物前来取食。比如上图中的火烈鸟,就用特殊的鸟喙滤取湖水中微小的卤虫。很可能在 2.9 亿年前的盐湖中,甲壳类和蜥形纲的前辈们就已经建立起类似的食物链了。图片来源自网络。
有许多中龙化石保留了腹部的内容物,从中可以清晰地分辨出臀头虾的残骸。如此看来,中龙略显恐怖的长牙其实主要用来捞取和咬碎这些小型甲壳动物。

一条保存非常完好的纳米比亚中龙前半身化石(A 图)。在它的肩胛骨下方有一个完整的大臀头虾(B 图箭头标示),那是它吞下的最后一口食物,可能是在掩埋的过程中从食道里挤出来的。图片来源自[1]。
除了臀头虾,研究者还在中龙的腹腔,粪便和呕吐物里发现了其他动物残骸——同类的幼体
。

Mangrullo 化石群发现的纳米比亚中龙化石,在腹腔中发现了食物残留的印痕(A)。放大图见 B 和 C,其中箭头标示的是臀头虾的外壳,锐角三角形指示的是中龙幼体的碎骨头。图片来源自[3] 。

一些在 Mangrullo 化石群发现的纳米比亚中龙粪化石(Coprolites)。A 图和 C 图中只有臀头虾的外骨骼;B 图中则有非常小的中龙幼体骨骼碎片(红线标示);D 图中不但有骨骼碎片,而且还有一枚牙齿(红色箭头标示)。图片来源自[1],标尺长度 1 厘米。

一些 Mangrullo 化石群发现的呕吐物化石(Regurgitalites),可以分辨出其中混杂着 中龙幼体的颈椎(cv),股骨(f),肋骨(fr),颅骨(fskb),牙齿(te)和其他骨骼(ub)的碎片。因为这里没有发现其他大型脊椎动物,所以可以基本确定来自中龙。看来它们会像许多现代动物一样,把不能消化,也不太好从肠道排除的硬质杂物呕吐出来。图片来源自[1],标尺长度 1 厘米。

另一处中龙化石发现点:巴西 Iratí化石群(位于大名鼎鼎的巴拉纳盆地)发现的中龙粪化石和肠内容物化石(Cololites)。A 图是一个粪堆。B 图粪里有一枚非常小的牙齿(标尺长度 1 厘米,放大后如 C 图,标尺长度 5 毫米)。D 图粪里有许多骨头碎片(箭头标示)。E 图是粪化石截面的 SEM 照片,可以看到许多臀头虾壳和中龙幼体的碎骨纠缠挤压在一起,这是中龙的两种主要食物。图片来源自[1]。
有人认为这是中龙同类相食的证据,也有人认为中龙会啃食同类的尸体。井蛙更倾向于前者。这么多的化石证据表明吃掉幼体的情况非常普遍,很难用食腐来解释。很可能因为 2 厘米的臀头虾对 1 米多长的中龙成体来说,还是太小了。中龙只能靠自己来填补食物链中间的空缺,在贫瘠的盐湖里建立起这样一条能量传递途径:藻类→臀头虾→中龙幼体→中龙亚成体→中龙成体。至于死去中龙的尸体,大概臀头虾会吃掉一部分,其他的中龙会吃掉一部分,剩下的残骸就交给微生物分解。腌在这种浓盐水里,尸体的腐烂速度会变得缓慢,就有更大的几率被湖底的沉积物覆盖掩埋。这也解释了为什么会有这么多保存完好的化石出土。
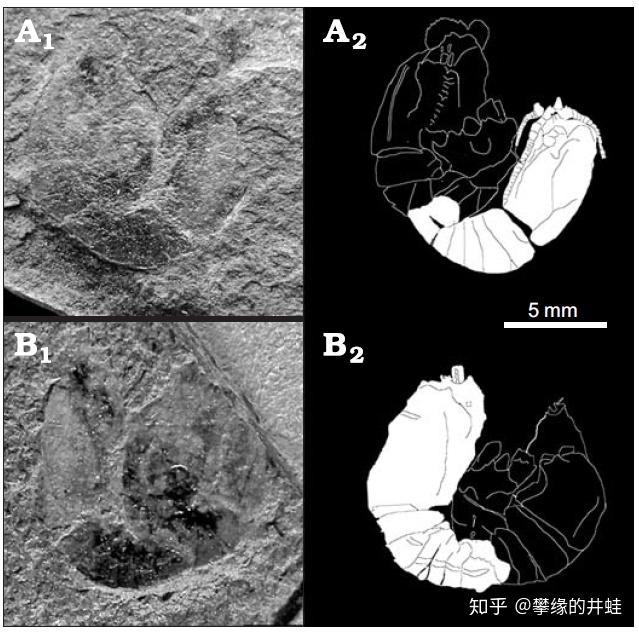
最为惊艳的化石要数中龙的胚胎。这也是人们发现的最早的羊膜动物胚胎化石。
Mangrullo 化石群发现的纳米比亚中龙化石。在成体腹部左侧有一个完整的胚胎骨骼(A 图)。B 图是胚胎和刚孵化幼体的体型对比。图片来源自[7]。
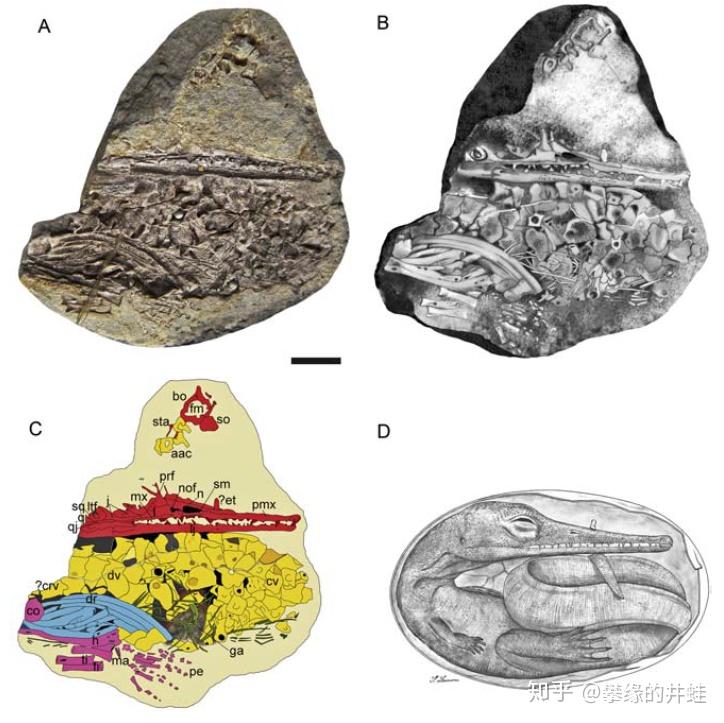
上图中的中龙胚胎骨骼结构和复原图。这个宝宝蜷缩成一团,但化石上没有卵壳的痕迹,它很可能是包裹在薄薄的卵膜里。图片来源自[7],标尺长度 5 毫米。
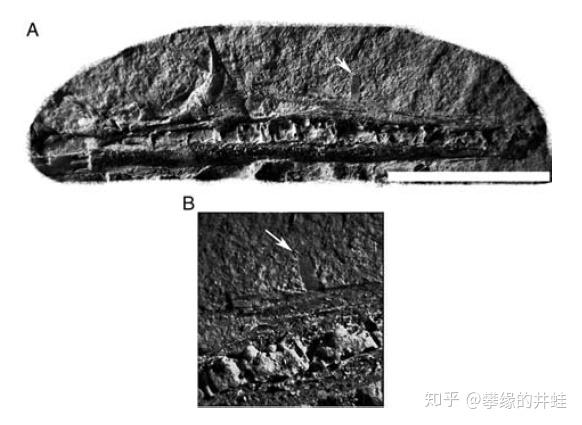
在这个胚胎的长嘴上发现了一个钉子状的小突起(箭头标示)。有人认为这是它的卵齿(egg tooth),用来顶破卵壳或卵膜的。图片来源自[7],标尺长度 1 厘米。
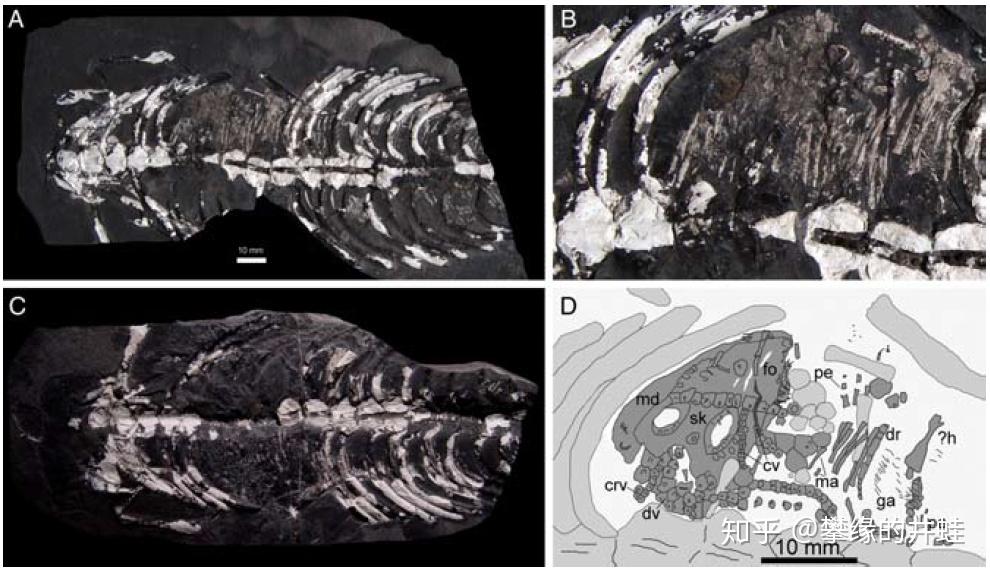
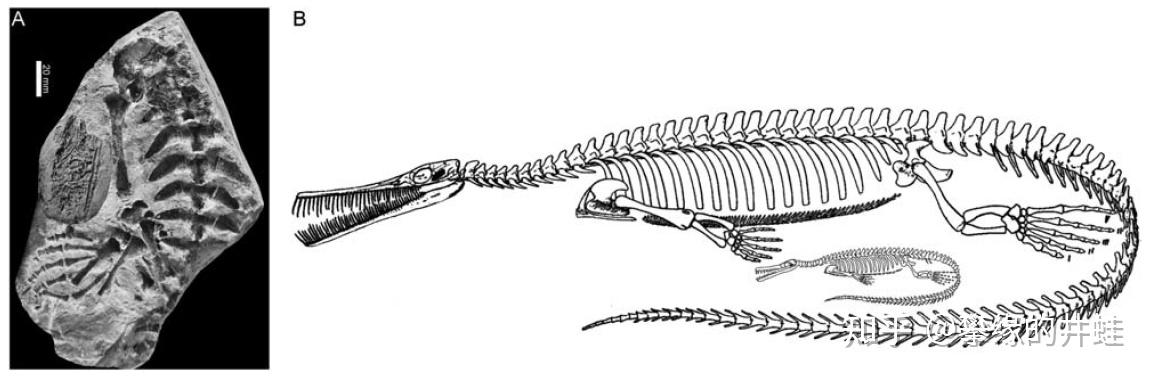
巴西 Iratí化石群发现的纳米比亚中龙化石。在它的腹腔里也有一个发育成型的胚胎(如 D 图所示)。图片来源自[7]。
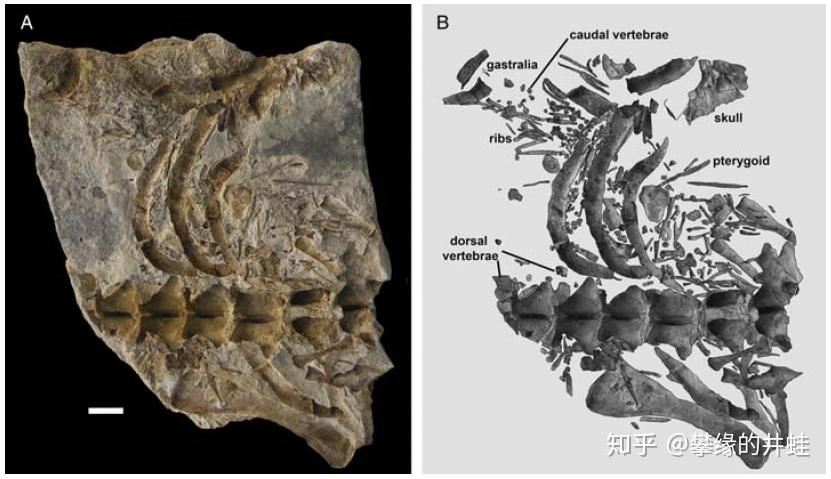

另外两块中龙化石。它们的腹腔里也发现了比较完整的幼体骨架(骨骼排列有序,基本可以排除是咬碎后吃掉的)。这些幼体比前面的胚胎大一些,可能更临近出生。图片来源自[7],标尺长度 1 厘米。
这些化石证据表明,中龙类的胚胎会在母体内充分发育,这也是对水栖生活的一种适应。它们要么孵化时间很短,说不定在水中产卵,幼体出生后马上用卵齿划开卵膜,开始自由生活。或者直接产下幼体(就像中生代的鱼龙那样)。如果是这样的话,那中龙确实没有理由再爬上岸去了,它们完全可以在水里度过一生。不过想一想,这么大的幼体,经过这么长时间的孕育,这么费力地生下来,结果有相当大的比例又被当成食物吃回去了。中龙的生存环境,还真是残酷呢。
说了分类地位和生活环境,还有猎食和繁殖行为,关于中龙类的简单介绍也差不多了。中龙类是羊膜类向水生生活进军的先锋探马。它们的生存年代之早,证明了羊膜动物演化出高度水栖的身体结构并不需要太长的时间。中龙类是幸运的,它们找到了鲨鱼和离片椎类统治的真空地带,在空寂的盐湖中建立起自己的统治。中龙类又是不幸的,它们的身体结构高度特化,生存空间被死死限定在狭小的盐湖里。当赖以生存的盐湖因为地质和气候变化最终消失,中龙便作为陪葬,一同掩埋在地层深处。在中龙消失 2500 万后,一场天地异变消灭了海洋河湖中的高级掠食者,觊觎这些生态位已久的羊膜类开始前赴后继的“下水”。“海爬”和“海兽”的时代正式开启,直到今天——当然,这是后话了。

游荡在二叠纪死海深处的一道魅影——中龙。图片来源见水印。
地球名片
生物分类:动物界-脊索动物门-脊椎动物亚门-四足形类-羊膜类-蜥形纲-副爬行动物-中龙类
存在时间:二叠纪早期
现存种类:无
化石种类:3 种
生活环境:内陆盐湖或浅海?
代表特征:高度水栖的体型,特化的长吻和牙齿
代表种类:纳米比亚中龙,凸方胸龙,圣保罗巴西龙
参考文献
[1] Rivaldo R. Silva, Jorge Ferigolo, Piotr Bajdek, et al., The Feeding Habits of Mesosauridae. Frontiers in Earth Science, Volume 5(2017), Article 23, doi: 10.3389/feart.2017.00023
[2] Dina Celeste Araújo-Barberena, Joffre Valmório de Lacerda Filho, Lílian de Lemos Timm, Mesossauro da Serra do Caiapó (Montividiu), GO. Sítios geológicos e paleontológicos do Brasil
[3] Piñeiro, G., Ramos, A., Goso, C., et al., 2012. Unusual environmental conditions preserve a Permian mesosaur−bearing Konservat−Lagerstätte from Uruguay. Acta Palaaeontologica Polonica 57 (2): 299–318
[4] Michel Laurin, Graciela H. Piñeiro, A Reassessment of the Taxonomic Position of Mesosaurs, and a Surprising Phylogeny of Early Amniotes. Frontiers in Earth Science, Volume 5 (2017), Article 88, doi: 10.3389/feart.2017.00088
[5] Graciela Piñeiro, Pablo Núñez Demarco, Melitta D. Meneghel, The ontogenetic transformation of the mesosaurid tarsus: a contribution to the origin of the primitive amniotic astragalus. PeerJ, (2016), DOI 10.7717/peerj.2036
[6] Graciela Piñeiro, Jorge Ferigolo, Alejandro Ramos, et al., Cranial morphology of the Early Permian mesosaurid Mesosaurus tenuidens and the evolution of the lower temporal fenestration reassessed. C. R. Palevol, 11 (2012), 379–391, doi: 10.1016/j.crpv.2012.02.001
[7] Graciela Piñeiro, Jorge Ferigolob, Melitta Meneghel, et al., The oldest known amniotic embryos suggest viviparity in mesosaurs. Historical Biology, iFirst article, 2012, 1–11












